Lipides
Catabolisme lipidique, ou lipolyse et β-oxydation, dégrade principalement les triglycérides en glycérol et acides gras, générant de l’ATP via l’oxydation mitochondriale lorsque le glucose est limité. Le glycérol entre dans la glycolyse, tandis que les acides gras subissent une activation, un transport et un clivage séquentiel jusqu’à former de l’acétyl-CoA, produisant du NADH et du FADH₂ pour la chaîne respiratoire.
Hydrolyse des triglycérides
Le catabolisme lipidique commence par l’hydrolyse enzymatique des triglycérides (TAGs) stockés dans le tissu adipeux par la lipase hormonosensible (HSL), activée en conditions de faible insuline et de forte glucagon, comme lors du jeûne. La HSL clive les TAGs en acides gras libres (AGL) et en glycérol, processus facilité par la monoacylglycérol lipase pour une dégradation complète. Les protéines perilipines présentes à la surface des gouttelettes lipidiques régulent l’accès des lipases, assurant une libération contrôlée des AGL dans la circulation sanguine, liées à l’albumine.
Activation et transport des acides gras
Les AGL sont activés dans le cytosol via l’acyl-CoA synthétase, consommant de l’ATP pour former l’acyl-CoA et de l’AMP + pyrophosphate (équivalent à deux ATP). Les acyl-CoA à longue chaîne ne traversent pas directement la membrane interne mitochondriale ; la carnitine palmitoyltransférase I (CPT1) située sur la membrane externe échange le CoA contre la carnitine, formant l’acyl-carnitine transportée par la carnitine-acylcarnitine translocase. La CPT2 régénère l’acyl-CoA dans la matrice mitochondriale. Le malonyl-CoA (issu de la synthèse des acides gras) inhibe CPT1 pour éviter un cycle futile.
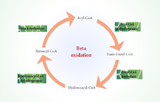
Voie de β-oxydation
Dans la matrice mitochondriale, la β-oxydation se déroule en quatre étapes répétitives par unité de deux carbones retirée :
(1) L’acyl-CoA déshydrogénase oxyde l’acyl-CoA en trans-Δ²-enoyl-CoA, produisant du FADH₂.
(2) L’enoyl-CoA hydratase ajoute de l’eau pour former le L-3-hydroxyacyl-CoA.
(3) La 3-hydroxyacyl-CoA déshydrogénase oxyde en 3-cétoacyl-CoA, générant du NADH.
(4) La thiolase clive avec le CoA pour produire de l’acétyl-CoA et un acyl-CoA raccourci.
Pour le palmitate (C16), sept cycles produisent huit molécules d’acétyl-CoA, sept NADH et sept FADH₂, générant ~106 ATP après le cycle de l’acide citrique et la phosphorylation oxydative.
Utilisation de l’acétyl-CoA et cétogenèse
L’acétyl-CoA entre dans le cycle de l’acide citrique, générant du NADH, du FADH₂ et du GTP pour la production d’ATP. L’excès d’acétyl-CoA en état de jeûne forme des corps cétoniques (acetoacétate, β-hydroxybutyrate, acétone) via la HMG-CoA synthase dans les mitochondries hépatiques, servant de carburant alternatif pour le cerveau et les muscles. La β-oxydation peroxysomale dégrade les acides gras à très longue chaîne (>C20), produisant du H₂O₂ au lieu du FADH₂.
Régulation et contextes physiologiques
Le catabolisme lipidique est régulé par les hormones : le glucagon et l’adrénaline/noradrénaline activent l’adénylate cyclase pour la phosphorylation de la HSL via l’AMPc, tandis que l’insuline inhibe par déphosphorylation. Le PPARα régule transcriptionnellement les gènes de la β-oxydation lors du jeûne ; l’AMPK inhibe l’ACC, réduisant le malonyl-CoA pour activer la CPT1. Des déficiences telles que CPT1/2 perturbent l’oxydation, entraînant une hypoglycémie hypokétotique.